Introducción
El Museo de La Plata (MLP) pertenece a la Facultad de Ciencias Naturales y Museo de la Universidad Nacional de La Plata (FCNyM-UNLP). Fue fundado en el año 1884 y posee uno de los patrimonios científicos y culturales más importantes de Latinoamérica, representado por sus colecciones provenientes de diferentes campos de las ciencias naturales y antropológicas. Su formación fue el producto de una intensa actividad científica, fundamentalmente desarrollada durante la segunda mitad del siglo XIX y principios del siglo XX (Farro, 2008). Las colecciones pertenecientes a la División Antropología (MLP-UNLP) constituyen un hito fundamental para la institución, ya que los primeros aportes configuraron las denominadas “Colecciones Fundacionales” donadas por Francisco Moreno en el año 1884. Entre la última década del siglo XIX y principios del siglo XX, las colecciones continuaron acrecentándose mediante materiales provenientes de las investigaciones de la propia institución, compras y donaciones (Del Papa, 2022). La guarda de restos humanos bajo la administración de una institución centenaria como el Museo de La Plata está atravesada por un complejo entramado en el que las formas de intervención -como prácticas curatoriales- estuvieron influenciadas por los marcos teóricos y alcances técnicos que imperaban en cada época. Desde su fundación como institución autónoma y su posterior integración a la Universidad Nacional de La Plata (Ricardi, 2014), la propia historia del museo ha dejado improntas en estas colecciones. Lo anterior da cuenta de las diferencias en los criterios de recuperación de los restos humanos como así también en los abordajes curatoriales, administrativos y de exhibición utilizados a lo largo del tiempo, que dieron lugar a la conformación de una de las mayores colecciones bioantropológicas. El imponente edificio que constituye el Museo de La Plata, de estilo neoclásico, fue diseñado en el año 1884 por el ingeniero alemán Carl Ludwig W. Heynemann y el arquitecto sueco Henrik A. Äberg. Cuenta con cuatro niveles: el subsuelo, que originalmente fue pensado para el alojamiento de diferentes talleres como así también una imprenta, mientras que la primera y la segunda planta fueron destinadas para la guarda y exhibición de diferentes colecciones; por último, el cuarto nivel lo constituye un ático donde se hayan locales de carácter secundario (Morosi, 2004).
La apertura de la totalidad del edificio y su inauguración oficial tuvieron lugar el día 19 de noviembre de 1889 (Morosi, 2004). El museo mantuvo activas modificaciones edilicias durante de todo el siglo XX. La planta del subsuelo sufrió cambios, ya que fue objeto de múltiples subdivisiones con la idea de facilitar nuevos espacios para laboratorios, como así también depósitos para el almacenamiento de colecciones. En este contexto, la División Antropología logró adquirir un conjunto de espacios dirigidos a la guarda de su acervo (Figura 1). La incorporación de nuevas dependencias durante la década de 1990, destinadas al funcionamiento de la Facultad de Ciencias Naturales y a nuevos laboratorios, permitió el traslado de estas actividades. Se desarrolló entonces un plan de intervención sobre el edificio histórico para salvaguardar la integridad edilicia y la reestructuración de las tareas dentro de la institución, fundamentalmente en el subsuelo.
Figura 1
Planta subsuelo edificio histórico del Museo de Ciencias Naturales. En verde, áreas para guarda del acervo de la División Antropología; en azul, emplazamiento de la cisterna; en rojo, ubicación de los restos óseos.
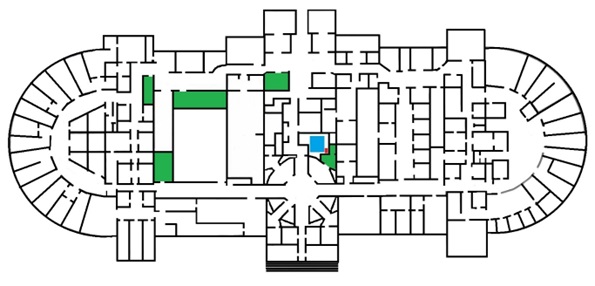
En este contexto es que, durante la remodelación en el sector correspondiente a la antigua cisterna, ubicada en la planta inferior (subsuelo) del museo (Figura 1), se realizó el hallazgo de restos óseos humanos, los cuales se encontraban fuera de los espacios asignados para su guarda, por lo que si iniciaron una serie de procedimientos orientados a dilucidar su origen.
De este modo, el objetivo de este trabajo es dar a conocer los resultados obtenidos en un caso de estudio en la aplicación del protocolo desarrollado por la División Antropología (FCNyM-UNLP) dirigido a la generación de información que permita aportar elementos para la contextualización de estos restos óseos humanos, mediante un abordaje multidisciplinario.
Protocolo de la División Antropología
En los primeros años del siglo XXI, nuevos paradigmas comienzan a cuestionar diferentes aspectos sobre tratamiento ético de los que deben ser objeto los restos humanos que se hallen bajo guarda institucional. Por este motivo, en el año 2006, desde la División Antropología (FCNyM-UNLP) se llevó a cabo un plan de gestión, dirigido fundamentalmente a desarrollar políticas curatoriales acordes a principios normativos delineados por el estado nacional (Ley Nacional 25.517) y principios éticos (UNESCO, 2005; ICOM, 2005).
Este protocolo (Expte. 1000_05286/2006 Actualización del Inventario y Documentación de las Colecciones de Restos Humanos de la División Antropología), que actúa como marco regulatorio de las actividades curatoriales de la División Antropología desde el año 2007 (ver Aranda et al., 2014; Castro et al., 2009; Del Papa y Pucciarelli, 2015; Del Papa et al.,2018) también ha podido ser replicado en otras instituciones (Del Papa, 2020). La metodología propuesta ha sido desarrollada en una serie de pasos procedimentales. La primera etapa está referida a aspectos técnicos curatoriales en las que se ha llevado a cabo el reacondicionamiento de las áreas de guarda (Castro et al., 2009). Una segunda etapa está dirigida a la conformación de la base de datos digitalizada que conforma el catálogo (Castro et al., 2009; Del Papa y Pucciarelli, 2015). Dadas la complejidad y las dimensiones que caracterizan a este acervo, y en virtud de que a lo largo del tiempo los criterios administrativos y curatoriales han variado sustancialmente (Del Papa et al., 2019), incluso considerando la guarda de restos humanos sin ingresar al catálogo, la aplicación de estos procedimientos debe contemplar pasos intermedios que permitan la mejor contextualización de aspectos tales como su origen geográfico y cronología (Del Papa et al.,2016) mediante la utilización de abordajes multidisciplinarios (Del Papa y Pucciarelli, 2015).
Caso de estudio
Los restos óseos humanos fueron encontrados detrás de la cisterna, en un rectángulo formado en dos de sus laterales por las paredes que separaban esta habitación de las contiguas, mientras que un tercer lateral era la estructura que sostenía la base de la cisterna (Figura 2), constituida por los mosaicos originales del edificio, donde no se encontró evidencia alguna de modificaciones.
Figura 2
Cuarto donde se hallaba la antigua cisterna. En rojo, lugar del hallazgo de los restos óseos. Nótese la presencia de una ventana que conecta con una de las áreas de guarda de la División Antropología.

Una vez realizado el hallazgo, la recuperación de los restos encontrados en el cuarto de la cisterna se llevó a cabo por integrantes profesionales de la División Antropología del Museo de Ciencias Naturales de La Plata. El procedimiento consistió en la utilización de técnicas de decapado mediante el uso de niveles horizontales sucesivos y arbitrarios de 10 cm. Se registraron los tipos de sedimentos constitutivos de la matriz que los contenían y la presencia de otros objetos.
Como resultado de las tareas de recuperación, se pudo constatar que la matriz en la que se hallaron los restos estaba formada principalmente por el material calcáreo usado como revoque de las paredes contiguas, junto con escombros de ladrillos. Asimismo, es importante notar que la matriz se hallaba dispuesta con diferencias granulométricas, con material más fino sobre la base de la estructura y que aumentaba de tamaño por encima del esqueleto. Posiblemente el esqueleto haya actuado como tamiz, permitiendo el paso de los granos más finos durante el lento proceso de depositación con el correr de los años. También fueron hallados diferentes elementos, posiblemente desechos de material en desuso (lapiceras, lámparas, botellas de vidrio, etc.), dispuestos por encima de los huesos. En virtud de encontrarnos ante restos óseos fuera de los espacios designados para el almacenamiento de la División Antropología y sin ningún tipo de identificación, se elaboraron dos hipótesis sobre su procedencia. La primera, la de encontrarnos frente a un caso forense, y la segunda, que los restos correspondieran a un individuo recuperado de un contexto arqueológico que, por motivos desconocidos, ha sido hallado en un área no designada para su guarda. Las autoridades del museo comenzaron con las diligencias administrativas mediante la apertura de un expediente durante el año 2007 (Expte. 1000-0080099/07-000- Hallazgos de un resto esqueletario en el sector del subsuelo del edificio del Museo de La Plata. Acompaña certificado de denuncia Policía Federal). En él se especifica que, enterados del hallazgo durante las obras, se realizó una denuncia ante autoridades de la Policía Federal, quienes recomendaron a esta institución tomar todas las acciones necesarias para esclarecer su origen, para lo cual solicitaban la intervención de la División Antropología.
Metodología
La utilización del protocolo fue llevada a cabo con el objetivo de generar toda aquella información que permita una adecuada contextualización de los restos humanos bajo guarda institucional, mediante el uso de múltiples líneas de abordaje. La generación de información en torno a diferentes características constitutivas de los restos humanos bajo guarda institucional (contexto) permite ampliar estos criterios y dar mayor relevancia a la colección (Castro et al., 2009). Por otro lado, los restos humanos ingresados (institucionalizados) poseen su propia biografía, una trayectoria con múltiples etapas y que, en el caso de restos humanos, comienza con el propio desarrollo ontogenético de los individuos y se extiende a momentos post mortem, que influyen en sus estructuras constitutivas y dejan evidencia de la interacción con su entorno. En este sentido, podemos agregar que, una vez ingresados como parte de una colección, los restos óseos también generarán una trayectoria fundamentalmente influida por políticas o enfoques curatoriales (Sardi y Del Papa, 2022).
Tomando en cuenta estas consideraciones, la primera etapa del protocolo fue evaluar potenciales agentes que podrían haber actuado en detrimento de la integridad de las unidades anatómicas (Del Papa et al., 2010). Para evaluar la integridad de los conjuntos anatómicos, se calculó el índice de fragmentación, de acuerdo con la propuesta de Mondini (2003), que utiliza la relación entre el número mínimo de elementos (NME) y el número mínimo de especímenes (NISP) (Lyman, 1984, 1994), cuya variación se encuentra entre 0 y 1. El grado de integridad observado en este sujeto limitó el uso de métodos para estimar la edad a la utilización de la propuesta original de Lovejoy et al. (1985) revisado por Buckberry y Chamberlain (2002) para la superficie auricular pélvica. El individuo analizado fue sexado mediante técnicas moleculares, siguiendo el protocolo descripto en Crespo et al. (2011). Se amplificó el gen de la amelogenina, de simple copia, localizado en la posición Yp11.2 del cromosoma Y, con su homólogo en la región Xp22.3-p22.1 del cromosoma X. Debido a que es dimórfico (su tamaño en el cromosoma Y es de 112 pares de bases y de 106 pares de bases para el cromosoma X), posee diferente cantidad de pares de bases en cada uno de los cromosomas en los que se encuentra, lo que permite diferenciar el sexo biológico del individuo.
Por otro lado, se evaluó el desgaste dental oclusal mediante la utilización de la metodología propuesta por Smith (1984), que considera la distribución de la pérdida de esmalte y dentina en las coronas dentales. Dicho método propone ocho estadios de desgaste, discriminando entre el patrón de dientes anteriores (incisivos y caninos), premolares y molares.
Con el objetivo de evaluar las modificaciones observadas en los especímenes en estudio, especialmente en contexto mortuorio, se llevó a cabo el relevamiento macroscópico de los siguientes agentes posdepositacionales: meteorización, acción de raíces, depositación química de carbonato de calcio (CaCO3) y óxido de manganeso (MnO2), fisura, fractura, pérdida de material óseo, marca de roedores, deformación plástica, aplastamiento y acción antrópica, específicamente presencia de pigmento y marcas de corte (Behrensmeyer, 1978; Binford,1981; Johnson y Haynes., 1985; Lyman y Fox, 1989; Villa y Mahieu, 1991; Lyman, 1994).
El análisis paleopatológico fue realizado a partir de una aproximación macroscópica mediante uso de lupa de 10X. Luego de la descripción de rasgos anómalos, se estableció una hipótesis diagnóstica para cada lesión (Campillo, 2001; Waldron, 2009). Para la evaluación de los cambios articulares degenerativos se tomó el modelo de Weiss (2006), clasificando las superficies articulares en grados 0 (ausente), 1 (leve), 2 (moderado) y 3 (severo). En el caso de la columna vertebral, se realizó un diagnóstico de dichos cambios para las articulaciones sinoviales interapofisarias y para las articulaciones entre cuerpos vertebrales adyacentes. Para el registro y clasificación de hernias discales, se siguieron las propuestas de Jiménez-Brobeil, Souich y Al Oumaoui (2009) y Üstündağ (2009). Las patologías de la cavidad oral se registraron y evaluaron de acuerdo con los criterios de Hillson (2000). Por último, se relevó la presencia de estrías y forámenes de vascularización como indicadores de formación de hueso nuevo en el periostio de cráneo y poscráneo (Weston, 2012), considerando la propuesta de Resnick (1985) para su descripción e interpretación.
Para el análisis del linaje materno se obtuvo una muestra de hueso largo, extraído del fémur derecho, utilizando un minitorno Dremel con disco para cortar hueso (aproximadamente 1 gramo de tejido óseo por extracción realizada). Las técnicas para la extracción de material genético consistieron en la descontaminación superficial, descalcificación y obtención del ADN antiguo mediante digestión enzimática y extracción con solventes orgánicos (Carnese, Mendisco y Keyser, 2010). Todos los trabajos de laboratorio, desde la preparación de la muestra, la extracción de ADN, como la tipificación de su linaje materno, fueron realizados en el laboratorio de Antropología Biológica del Centro de Estudios Biomédicos, Básicos, Aplicados y Desarrollo (CEBBAD) Universidad Maimónides. La secuenciación del fragmento de ADN mitocondrial amplificado fue realizada en la unidad de genómica del Instituto Nacional de Tecnología Agropecuaria (INTA). El ADN fue purificado con columnas de sílica y se amplificó por PCR la Región Hipervariable 1 (RHV-1) del genoma mitocondrial (ADNmt), utilizando los cebadores y el protocolo descripto en Carnese et al. (2010) y Dejean et al. (2011). El ADNmt amplificado fue secuenciado sobre la base de los mismos cebadores descriptos en Carnese et al. (2010). Se consideraron válidos los resultados al obtener secuencias coincidentes a partir de dos amplificaciones.
Las secuencias obtenidas se compararon con los perfiles de los operadores que realizaron los trabajos moleculares (CMC y CBD), para descartar contaminaciones. Las secuencias fueron revisadas y alineadas con el programa MEGA 7 (Kuma, Stecher y Tamura, 2016) y la secuencia consenso obtenida fue comparada con la Secuencia de Referencia de Cambridge Revisada -SRCr- (Andrews et al., 1999). La identificación del haplotipo del individuo se realizó con el software Haplogrep 2 (Weissensteiner et al., 2016), basado en Phylotree 17 (van Oven y Kayser, 2008). Se contrastó con una base de datos de ADNmt de poblaciones antiguas, históricas y actuales obtenida por revisión bibliográfica y que cumplieran con el requisito de tener la RHV-1 completa. Solo se emplearon en el análisis aquellas que pertenecían exclusivamente al clado del individuo analizado, ya que el objetivo es contextualizar su posición dentro de la filogenia de su linaje y, de esta manera, inferir el posible origen geográfico materno. Por este motivo, las secuencias antiguas se agruparon según procedencia geográfica, mientras que para las secuencias de poblaciones nativas americanas actuales se respetó el criterio de adscripción con el cual fueron presentadas en cada una de las publicaciones originales (Ginther et al., 1993; Dejean et al., 2008; Moraga et al., 2010; De Saint Pierre et al.,2012; De la Fuente et al., 2015; Raghavan et al., 2015; Arencibia et al., 2019; Parolin et al., 2019; Motti et al., 2020; Nakatsuka et al., 2020; Postillone et al., 2020a, 2020b).
Se construyó una red de haplotipos para el linaje de la muestra con el programa Network v.5.0 (Bandelt, Forster, y Röhl., 1999; Fluxus-enineering.com). Los pesos evolutivos para cada sitio polimórfico se asignaron según los criterios descriptos en Crespo et al. (2020) con el fin de minimizar las reticulaciones y artefactos en la red. Finalmente, se utilizó el cálculo de posprocesamiento con el algoritmo de máxima parsimonia de Steiner para simplificar la red (Polzin y Daneschmand, 2003). Por último, la contextualización cronológica fue realizada en el Laboratorio de Tritio y Radiocarbono (LATYR-CIG-CONICET-UNLP) mediante un fechado radiocarbónico sobre fragmento del fémur derecho, a partir de la calibración correspondiente para el hemisferio sur (SHCal20) (Hogg et al., 2020, Programa Oxcal 4.4).
Resultados
Para la determinación de edad, la suma de los valores de las variables observadas en la superficie auricular fue igual a 15. Este valor corresponde al estado VI de desarrollo, que le asigna a este individuo al grupo etario de adulto mayor. En referencia a la determinación del sexo, a partir de la amplificación del gen de la amelogenina se pudo comprobar la presencia de una banda de 106 pares de bases, que indican que el sexo biológico del individuo analizado es femenino.
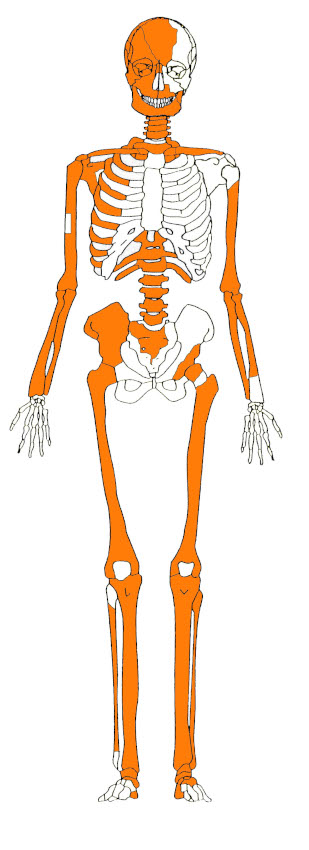
Los diferentes índices de abundancia permitieron establecer un NISP total de 83 especímenes (NISP craneal, 3; NISP poscraneal, 80). El individuo recuperado tiene una representatividad del 37% del esqueleto (Figura 3). El estado de preservación de las unidades anatómicas posee un IF de 0,33, que indica una mala conservación (Mondini, 2003).
Figura 3
Unidades anatómicas presentes.
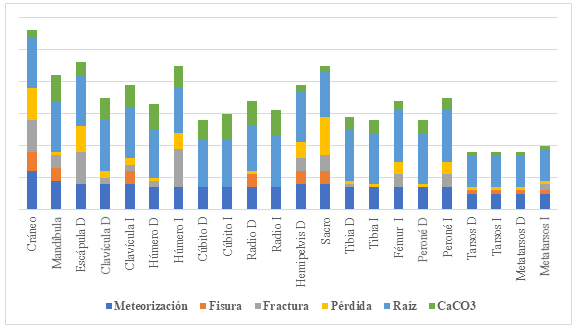
De la observación macroscópica de las diferentes unidades anatómicas se desprende que ha actuado un conjunto de agentes posdepositacionales en la modificación y posterior pérdida de material. En primer lugar, y con mayor intensidad y frecuencia, están las marcas de raíces: han afectado entre el 50% y 80% de la superficie de las unidades anatómicas analizadas. El segundo agente posdepositacional en grado de impacto fue la meteorización. Este agente se define por la destrucción de los componentes orgánicos e inorgánicos microscópicos originales del hueso, que son separados unos de otros y destruidos por los agentes físicos y químicos que actúan en el hueso in situ, en la superficie o dentro de la zona del suelo. En cuanto al cráneo, el 60% de la superficie presenta un estadio de meteorización de grado 3, caracterizado por sectores de hueso compacto áspero, resultando en una textura fibrosa que afecta a los primeros 1,5 mm de profundidad con fracturas de bordes redondeados (sensuBehrensmeyer, 1978). Esto demuestra que este elemento óseo ha estado expuesto durante un período prolongado a las fuerzas modificadoras de la meteorización. A diferencia del cráneo, el esqueleto poscraneal evidencia un grado de meteorización 1, donde la superficie ósea no presenta signos de agrietamientos o exfoliación. Otro agente identificado es la depositación de carbonato de calcio. Esto se correlaciona con el contenido de carbonatos del sedimento que rodea a los huesos, que puede variar a lo largo del tiempo por lixiviación y pedogénesis, entre otros factores. Por último, la fisura, fractura y pérdida se expresan en valores relativamente bajos o no fueron registrados durante el relevamiento (Figura 4).
Figura 4
Porcentajes de presencia de agentes tafonómicos para cada unidad anatómica en estudio. Nota: CaCo3= Carbonato de calcio.
Los análisis paleopatológicos permitieron observar un desgaste alto, que alcanzó en general un grado 6 en incisivos y premolares. En los molares de la arcada superior, el grado alcanzado fue de 8, con una separación de la porción de la corona correspondiente a cada raíz del molar. Los signos patológicos más frecuentes fueron aquellos vinculados con la cavidad oral y con la degeneración articular. Entre las del primer grupo se encuentra la pérdida de hueso alveolar de tipo horizontal o periodontitis, que afectó a la totalidad de ambas arcadas. Las patologías infecciosas en esta cavidad se manifestaron como lesiones periapicales en los dientes canino y 2º incisivo derechos e izquierdos y 2º premolar izquierdo, todos pertenecientes a la mandíbula maxilar inferior (Figura 5a).
Figura 5
Patologías descriptas. Nota: A = abscesos periapicales; B y C = osteoartritis (OA); D = eburnación; E = osteoartritis (OA); F = nódulo de Schmörl ; G = osteoartritis (OA).
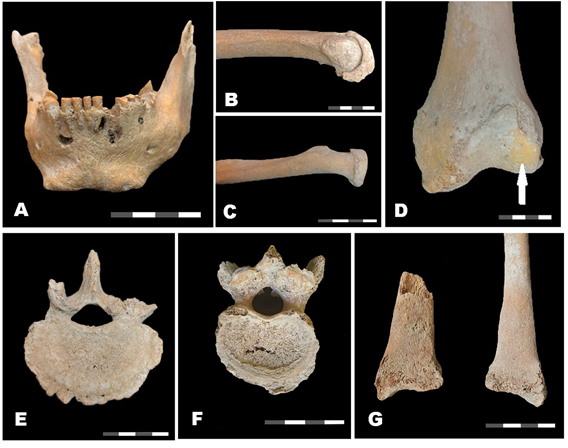
En cuanto a las patologías osteoarticulares, se reconocieron signos consistentes con enfermedad degenerativa articular en varias unidades anatómicas. En las carillas proximales del húmero derecho y radio izquierdo se registraron señales moderadas, mientras que en la epífisis proximal de dicho húmero y en la epífisis distal del cúbito izquierdo se observaron las mismas modificaciones, aunque de manera severa (Figura 5b y 5c). En la articulación de la rodilla izquierda también se registró de esta forma, con presencia de eburnación en el cóndilo externo femoral (Figura 5d), al igual que los calcáneos y astrágalos de ambos pies. En la columna vertebral, cambios degenerativos fueron relevados en todos los elementos, que son predominantemente leves en la región cervical y torácica alta y disminuyen en la torácica baja y lumbar (Figura 5e) y sacra. No se observó eburnación en ninguna de las articulaciones vertebrales, a la vez que fue característica en las articulaciones interapofisarias la presencia de osteofitosis, alteración del contorno, sumando a estos rasgos la porosidad en el caso de las articulaciones entre cuerpos vertebrales. También se relevó una lesión atribuible a una hernia discal vertical, en forma de nódulo de Schmörl, localizada en posición central sobre la cara inferior de la vértebra torácica 10 con una severidad de grado 1 o leve, de forma lineal con dimensiones de 13,9 × 1,8 mm (Figura 5f). Estas anormalidades degenerativas son el resultado del estrés funcional por tracciones capsulares sistemáticas en individuos mayores de 50 años (Luna et al., 2017).
Otra característica importante fueron las posibles señales de enfermedades infecciosas inespecíficas en el fragmento distal del radio derecho. En la cara anterior del tercio distal de su diáfisis fue relevada una lesión focal, que comprometía aproximadamente un 10% del hueso, en forma de hueso lamelar delimitando estriaciones y pequeños forámenes, en forma de una simple capa de hueso nuevo subperióstico (Figura 5g). Por último, cabe destacar que no se observaron signos de traumas ni de patologías metabólicas.
Se logró obtener el linaje materno del individuo a partir de la amplificación de la RHV-1 del ADNmt, con una secuencia que presentaba las siguientes mutaciones en comparación con la SRCr: 16150T - 16177G - 16187T - 16189C - 16223T - 16325C - 16362C, pasible de ser adscrito al clado D1g, el cual es estrictamente patagónico (Bodner et al., 2012). El haplotipo del individuo analizado posee tres mutaciones adicionales en relación con el nodal de su clado (D1g + 16150T, 16177G y 16189C). En la Figura 6 se presenta la red de haplotipos de todos los linajes D1 reunidos en la base de datos a partir de muestras de poblaciones actuales, históricas y antiguas de Patagonia.
Tabla 1
Muestras antiguas y nativas actuales utilizadas para realizar la red de haplotipos del clado D1g. En la primera columna: 1: muestras arqueológicas; 2: muestras actuales.
|
|
Región/población |
n |
Muestras utilizadas |
Referencias |
|
LP Sample |
1 |
LP |
Este trabajo |
|
1 |
Patagonia septentrional |
8 |
FD6, FD13, MB34, MB12, MB48, Ch1, PA8 y PA10 |
Crespo et al. (2017), Postillone et al., (2020a, 2020b) |
|
Patagonia meridional |
9 |
LS1, LS2, LS7, LS12, LS21, LS27, LS31, La Azucena y Cerro Johnny |
Arencibia et al., (2019), Parolin et al. (2019), Nakatsuka et al., (2020) |
|
Norte de Tierra del Fuego |
13 |
Faro Méndez, Río Grande, Pozo, MA577, QQN20, QQN31, QQN32, QQN34, QQN39, QQN 40, QQN43, QQN51 y QQN53 |
Raghavan et al. (2015), Motti et al. (2020), Nakatsuka et al. (2020) |
|
Península Mitre e Isla de los Estados |
6 |
PZ7, PZ8, Caleta Falsa 3, Caleta Falsa 7, Caleta Falsa 8 y Policarpo |
Crespo et al. (2020), Nakatsuka et al. (2020) |
|
Archipiélagos occidentales |
2 |
IPK03, IPK11 |
De la Fuente et al. (2015) |
|
Canal Beagle |
4 |
PZ3, PZ4, CBD12590, MG30b |
Dejean et al. (2008), Crespo et al. (2020), De la Fuente et al. (2015) |
|
2 |
Patagonia septentrional |
12 |
(Mapuche Isla Huapi) Hu09, 01, 07 (n = 3), 04, 02, 05 (n = 3), 08 (n = 2) |
Moraga et al. (2010) |
|
10 |
(Mapuche Anecón Grande) Anecón 11 (n = 4), Anecón 12 (n = 4), Anecón 13, Anecón 14 |
Ginther et al. (1993) |
|
3 |
(Mapuche Temuco) Ma18, 24, 29 |
De Saint Pierre et al. (2012) |
|
15 |
(Mapuche Lonco Vaca) Marg100, 127, 71, 83, 151, 137, 113, 152, 102, 133, 90, 39, 99, 110, 44 |
De Saint Pierre et al. (2012) |
|
24 |
(Huilliche San Juan) Hui80, 34, 92, 41, 123, 86, 02, 115, 105, 01, 09, 13, 04, 27, 07, 54, 32, 68, 98, 11, 15, 61, 73, 37 |
De Saint Pierre et al. (2012) |
|
16 |
(Pehuenche) T88, 19, 83, 24, 27, 44, 48, 58, 78, 81, 99, 09, 63, 94, 33, 72 |
De Saint Pierre et al. (2012) |
|
9 |
(Tehuelche) Tehlr02, 06, 08, 04, 03, Teh 56, 50, 13, 26 |
De Saint Pierre et al. (2012) |
|
Archipiélagos occidentales |
1 |
(Kaweskar ) K01 |
De Saint Pierre et al. (2012) |
|
Canal Beagle |
9 |
(Yámana) Ya02, 20, 05, 08, 14, 19, 24, 25, 09 |
De Saint Pierre et al. (2012) |
|
|
Total |
142 |
|
Figura 6
Red de haplotipos correspondiente a los linajes D1 presentes en poblaciones actuales, históricas y antiguas de Patagonia.
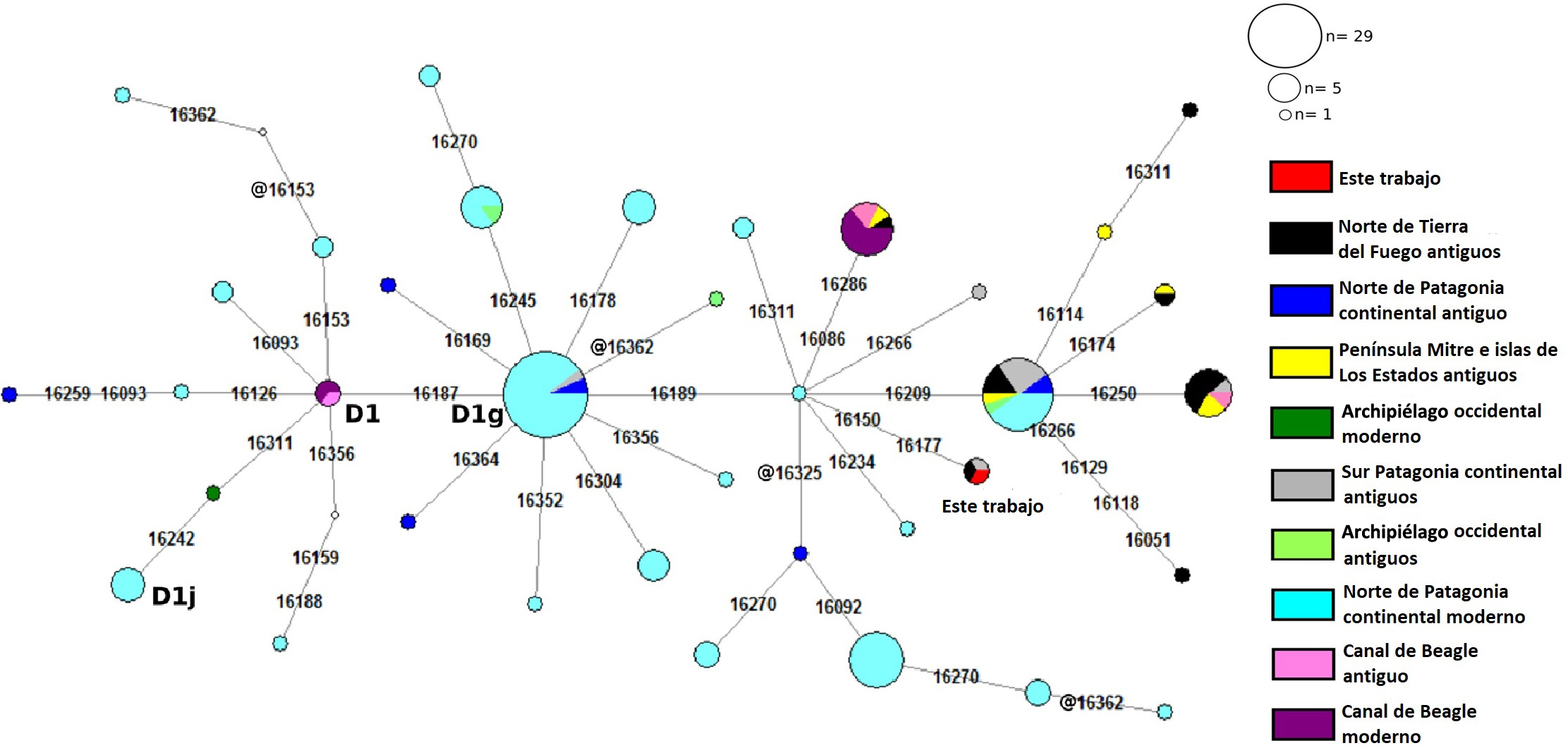
Por último, el análisis radiocarbónico determinó una cronología de 2420 ± 60 años AP (LP-1920; hueso humano, 687 años cal AC-137 años cal AC)
Discusión y conclusiones
La aplicación de este protocolo permitió generar un conjunto de información en torno a diferentes aspectos de la biografía de los restos óseos humanos objeto de este análisis. Se trata de un individuo adulto mayor de sexo femenino, cuya asignación cronológica corresponde al Holoceno tardío inicial, que probablemente estaba en una posición subsuperficial al momento de su recuperación en el campo. Esto estaría evidenciado por la fuerte acción de las raíces en la superficie cortical y la actividad de diferentes agentes atmosféricos que deterioraron, por meteorización, la mayoría de las unidades anatómicas. Se logró la detección de patologías infecciosas en la cavidad oral y otras patologías osteoarticulares degenerativas, congruentes al grupo etario adulto mayor al que fue asignado el individuo. Las patologías osteoarticulares degenerativas no presentaron rasgos particulares en su prevalencia, distribución y severidad, por lo cual no fue posible establecer su relación con algún tipo de actividad física. Similar es el caso de la reacción perióstica observada, cuyo carácter leve y focal imposibilita la realización de una hipótesis diagnóstica.
Un resultado por demás significativo fue la posibilidad de haber obtenido información sobre el sexo biológico (femenino) y el ADNmt de este individuo, perteneciente al subclado D1g. Este linaje fue identificado por primera vez en poblaciones actuales de la comunidad mapuche de Anecón Grande (Departamento de 25 de Mayo, provincia de Río Negro, Argentina), durante los trabajos bioantropológicos y genéticos pioneros realizados por el Dr. Raúl Carnese (Ginther et al., 1993). También se detectó en poblaciones cosmopolitas de distintas localidades del centro de Chile descriptas en Horai et al. (1993). A partir del estudio de genomas mitocondriales completos, Bodner et al. (2012) describieron y dieron a conocer la variabilidad del linaje D1g. Este haplotipo no se ha encontrado en ninguna de las poblaciones nativas actuales de la Patagonia, pero sí ha sido descripto en dos individuos antiguos analizados a partir de estudios de ADNa. El primero de ellos, publicado por Arencibia et al. (2019), pertenece al individuo SAC 22-1-1 procedente de la región del lago Salitroso, en el noroeste de la provincia de Santa Cruz (Argentina), datado en 704 ± 42 años C 14 AP y determinado como un adulto de sexo masculino (García Guraieb et al., 2015). El segundo individuo que comparte este linaje, denominado “Río Grande” y fechado en 520 ± 30 años 14C AP, fue presentado por Nakatsuka et al. (2020). A partir de estos resultados, es posible inferir que el individuo analizado tendría una ancestría materna oriunda de la Patagonia, y que compartiría linaje con individuos que habitaron su sector meridional.
En la actualidad, dicho clado tiene una presencia importante en poblaciones del Cono Sur de Sudamérica, especialmente en Patagonia, región en la cual alcanza sus mayores frecuencias y posee su mayor diversidad. Por estos motivos, Bodner et al. (2012) proponen a la región patagónica como su lugar de origen y, según los tiempos de coalescencia para este linaje, le adscriben una temporalidad cercana al momento de ingreso de las primeras poblaciones humanas.
Todas las acciones orientadas hacia la contextualización, fundamentalmente sobre su probable origen geográfico y su posición cronológica, permiten afirmar que estos restos humanos pertenecen a un conjunto proveniente de un contexto arqueológico enviado al museo para su guarda respecto del cual se desconocen los detalles históricos o la razón por la que fueron depositados en el lugar donde se los halló.
El presente trabajo y los resultados obtenidos en la aplicación del protocolo permiten dar resignificación científica, revalorizando el aporte realizado desde diferentes líneas de abordaje que permitieron una aproximación al estilo de vida de este sujeto y a la reconstrucción de los hechos.